In allen Lehrbüchern kann man nachlesen, dass der erste entdeckte morphogenetische Gradient dadurch gebildet wird, dass das so genannte bicoid-Protein am vorderen Pol des Embryos der Taufliege Drosophila synthetisiert wird und von dort durch Diffusion nach hinten gelangt. Dieses Dogma hat jetzt ein internationales Wissenschaftlerteam in der aktuellen Ausgabe der Fachzeitschrift „Development“ widerlegt.
Morphogenetische Gradienten bilden ein zentrales Konzept der Entwicklungsbiologie, dessen Ursprung mehr als hundert Jahre zurückreicht. Heute werden sie definiert als Substanzen, deren Konzentration mit zunehmender Distanz von einem Referenzpunkt oder einer Referenzlinie monoton abfällt und die, entsprechend ihrer Konzentration, verschiedene Gene aktivieren. Ein morphogenetischer Gradient legt daher im Embryo die Position entlang einer Achse fest.
Forschungsobjekt Drosophila
Zum ersten Mal wurde ein morphogenetischer Gradient 1986 nachgewiesen, und zwar für das bicoid-Genprodukt, dessen Konzentration mit der Distanz vom vorderen Pol eines Drosophila-Embryos abnimmt und so im wesentlichen das Schicksal der Zellen bestimmt, die Kopf und Thorax der Drosophila Larve bilden. Damals postulierten Hans Georg Frohnhöfer und Christiane Nüsslein-Volhard vom Max-Planck-Institut in Tübingen aufgrund genetischer und entwicklungsbiologischer Experimente im frühen Embryo einen Gradienten des bicoid-Genprodukts.
Gleichzeitig gelang im Labor von Professor Markus Noll vom Institut für Molekularbiologie der Universität Zürich der erste Nachweis des postulierten Gradienten und zwar in Form von bicoid-mRNA, die im Mikroskop von Stefan Baumgartner, seinem damaligen Doktoranden, im Embryo sichtbar gemacht wurde. Nolls Gruppe zeigte damals auch, dass diese mRNA für ein Protein kodiert, das andere Gene reguliert. Damit war offensichtlich, wie das Produkt des bicoid-Gens als Morphogen wirkt, nämlich indem es in unterschiedlichen Konzentrationen verschiedene Gene aktiviert.
bicoid-Protein im Embryo nachgewiesen
Zwei Jahre später, 1988, haben Wolfgang Driever und Nüsslein-Volhard dann einen Gradienten für das bicoid-Protein im Embryo nachgewiesen. Dabei postulierten sie, dass die für das bicoid-Protein kodierende mRNA strikt am vorderen Pol des Embryos lokalisiert bleibt, obwohl dies einer zwei Jahre zuvor von Nolls Team publizierten Evidenz widersprach.
Nach der Vorstellung von Driever und Nüsslein-Volhard entstand der Gradient des bicoid-Proteins dadurch, dass die am vorderen Pol lokalisierte bicoid-mRNA in Protein übersetzt wurde, das nach hinten diffundierte und so ein Konzentrationsgefälle bildete. Dieses Modell wurde seither in allen entwicklungsbiologischen Lehrbüchern wie ein Dogma gelehrt.
{2l}
Modell endgültig widerlegt
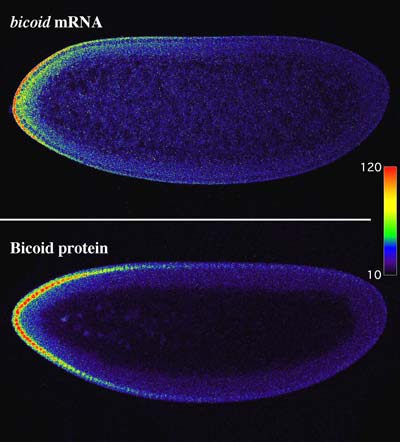
In „Development“ haben nun die Labors von Baumgartner und Noll mit den heutigen modernen Methoden ihre Befunde von 1986 bestätigt und zusätzlich gezeigt, dass der Gradient der bicoid-mRNA mit demjenigen des bicoid-Proteins so gut wie identisch ist. Damit wurde das von Driever und Nüsslein-Volhard postulierte und in Lehrbüchern reproduzierte Modell endgültig widerlegt.
Der morphogenetische Gradient des bicoid-Proteins bildet sich nicht durch Diffusion des ausschliesslich am vorderen Pol synthetisierten bicoid-Proteins, sondern einfach durch die Synthese des bicoid-Proteins auf der bicoid-mRNA, die dabei als Matrize dient und einen Gradienten bildet.
Nun stellt sich die Frage, wie der bicoid-mRNA-Gradient zustande kommt, da im unbefruchteten Ei die bicoid-mRNA am vorderen Pol lokalisiert ist, wie ebenfalls 1986 in der Publikation von Nolls Gruppe zum ersten Mal gezeigt wurde.
Die Forscher schlagen ein Modell vor, wie dies geschieht: Die bicoid-mRNA wird nach der Befruchtung vom vorderen Pol losgelöst und mit Hilfe des als Staufen bezeichneten Proteins und bekannten Proteinmotoren entlang einem Netzwerk aus Microtubuli in einer dünnen Schicht direkt unter der Oberfläche des Embryos vom vorderen Pol nach hinten transportiert. Da aber die Orientierung der Microtubuli zufällig ist, wird die mRNA, ähnlich wie bei der Diffusion, nur wegen ihres Konzentrationsgefälles langsam nach hinten transportiert (etwa hundert Mal langsamer als bei einem gerichteten Transport entlang Microtubuli).
Funktion des Microtubuli-Netzwerks enthüllt
Dieses Modell erklärt auch zum ersten Mal eine Funktion dieses Microtubuli-Netzwerks, nämlich die Bildung des bicoid-mRNA- Gradienten. Sobald die endgültige Form des mRNA-Gradienten erreicht ist, bricht dieses Netzwerk zusammen. Dies geschieht zu Beginn des so genannten synzytialen Blastoderms, wenn die Zellkerne nach 9 Kernteilungen im Innern des Embryos die Oberfläche erreichen.
Der bicoid-Gradient entsteht also weder durch Diffusion noch durch das Erreichen eines stationären Zustands, sondern sein Aufbau ist zeitlich perfekt mit der Entwicklung des Embryos synchronisiert. Daher erreicht er seine endgültige Form, wenn das bicoid-Protein, entsprechend seiner Konzentration, verschiedene Gene in den Zellkernen aktivieren muss.
(idw – Universität Zürich, 28.01.2009 – DLO)